
The Evolution of Human Intelligence and Neural InhibitionAdam Kircher and Karen K. Glendenning |
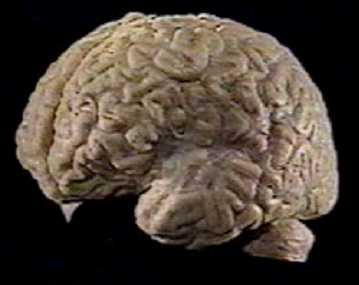
The left cerebral hemisphere of the human brain. The outer layers of the hemispheres are the neocortex which is the characteristic of humans that seems to underlie our uniqueness. Yet the makeup and connectivity of the cortex is similar in all vertebrate animals. |
When we contemplate the extraordinary abilities and accomplishments of humans, we are usually most impressed with our intelligence. We feel we are unique creatures on this planet. And the source of our uniqueness lies in our intellectual capacities. Are we really so unique?
Many hypotheses have been put forward as to what makes us unique. However, most of these have had be discarded after similar functions have been found in animals. Some of these theories were: tool use, tool making, tradition, innovation, trade, barter, playing games, morality. Yet, all of these behaviors have been found in other species. It has also been thought that it is the adoption of symbolic cognitive processes that gave our kind the final edge in natural selection. But these have also been found in other species birds as well as apes! Even our ability to attribute a mental state to others does not seem to be uniquely human since finding behaviors indicating deceitfulness in the great apes. One negative trait that may be unique to humans is genocide. We are the only species with weapons of mass destruction!
However, even this behavior depends on our intelligence. Intelligence seems to underlie whatever uniqueness we may have. Therefore, this is the characteristic to which we have turned our attention. Intelligence is a biological phenomenon and as such can be looked at from a comparative or evolutionary point of view. That is, there has been an element of inevitability in the process by which we have become what we are today. And, like other biological processes, intelligence has been and continues to be pressured by the forces of natural selection. The human brain has been honed for the last 10 million yearsbefore, in fact, there were humans! Perhaps by looking at the selective pressures on our brain we can determine the purpose of the enhanced capacity we seem to possess.
That is, there appears to be no correlation whatever between the achievement in the human lineage of behavioral modernity and anatomical modernity. That is, the emergence of anatomically modern Homo sapiens would have considerably predated the arrival of behaviorally modern humans. But these behavioral innovations would have been established within a preexisting species. Unfortunately, behaviors and brains not preserved only skulls. We cannot examine or test the brains of a Neanderthal or Cro-Magnon man. We only have their bones. It is for that reason, this work is particularly important, for although we cannot look at the brains of our forebearers, we can look at the brains of other species which are representative of stages in mans evolution.
The arrival of the modern cognitive capacity did not simply involve adding just a bit more neural material, nor did it involve adding any major new brain structures. The basic brain design remains remarkably uniform among all the higher primates and as this work shows among all mammals.
While there is an evolutionary continuum of variation, there is also general agreement that a prime result of evolutionary pressure has been the selection for human intelligence. Perhaps the main biological correlate of the intelligence of humans is with the increase in size of the neocortex. However, we propose here that, in addition to the size of neocortex, the amount of neural inhibition to which the cortex is subjected may play a major role in human intelligence. That is, an expanded brain that is out of control is not helpful! The cortex is totally dependent on input from the thalamus. This is true in our brain and also in all vertebrate animals. The thalamus determines what the cortex will see, hear, or smell. And in turn, after the cortex has made a decision on how to act on the information, the thalamus redirects the information to other centers that allow us to run, speak, eat etc.
There must be modulation of this enhanced cortex. Our work shows a primary source of this modulation the enhanced inhibitory capabilities of the thalamus. By its influence on our neocortex, there is greater control of neural processing. We propose that it may be our ability to inhibit our cortex that has resulted in our increased intelligence. But this change appears to be present in all vertebrates, mammals and nonmammals.
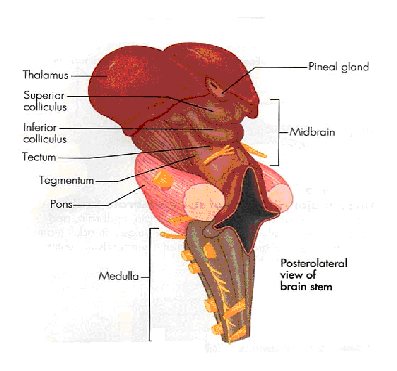 |
The thalamus is at the most forward part of the brainstem and channels all of the information going to and returning from the cerebral cortex. Since the source of inhibitory input to the cortex is from the thalamus, an examination of this structure alone could be predictive of cortical function. We dont need to look at the cortex to tell who is bright! |
A problem with evolutionary analysis is that it will never be an exact science. One reason for this is that our preconceptions may mislead us as to what is the objective reality. However, by quantitative measurements of the particular neurotransmitters in the thalamus, preconceptions are minimized. We examined the neurotransmitters in the thalamus of a series of species representative of our ancestral heritage,
Because there are close parallels in the evolutionary development of dorsal thalamus and cerebral neocortex, a difference in the thalamus in mammals (see hypothetical ancestor '1' in Figure below) and their premammalian ancestors ('0' in Figure below) might well be a concomitant of the enhanced cortical development of the cerebrum seen in the mammals over the same period of geological time -- perhaps a demand placed on the dorsal thalamus by the emergence of mammalian cortex itself.
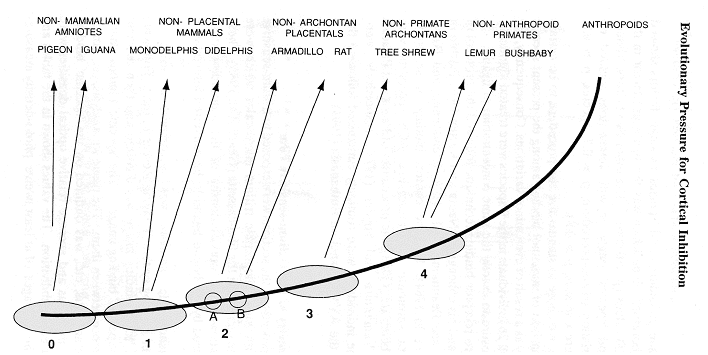
By
an examination of diverse species chosen to represent a phylogenetic series
leading to man, we can show one step in the consequences of selective pressure
on the mechanisms of brain function. Our conclusion is that this
step, an increase in the potential for inhibition of the cortex, is probably
a major component of the evolution of human intelligence. In the present
study, the idea of an increasing amount of GABA inhibition over the Anthropoid
ancestral lineage was investigated using seven mammals and two non-mammals.
These species were selected to yield at least 5 differing levels of recency
of last common ancestor with Anthropoids. That is, they show something
like what we were like at different stages of our evolution.
Thus, any feature found in common among these widely different species
would most likely have also been present in our most ancient ancestor.
We have started a study to examine experimentally
the idea of an increasing amount of GABA inhibition over the anthropoid
ancestral lineage. Our investigation used seven mammals and two non-mammals
selected to show what we were like at different stages of our evolution.
The features found in common among these different species would most likely
have also been present in our most ancient ancestor. Each of these
species was subjected to exactly the same laboratory procedures which determine
the amount of the inhibitory neurotransmitter GABA present in the thalamus.
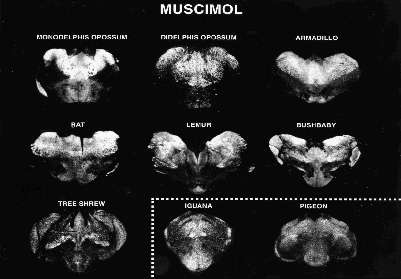
Prints of images made directly from x-ray films of the relative degree of attachment of the neurotransmitter GABA (as shown by muscimol label . Each section of the brain thalamus is taken through the thalamus. The areas of densest binding appear white and areas of little or no binding appear black. Even though there are seven mammals and two non-mammals the profile of bound GABA is very consistent with most being near the top (dorsal) and less in the bottom (ventral) subdivisions. (Two subdivisions are difficult to see: the actual pattern for all 4 subdivisions is, low-high-low-high, dorsal to ventral). |
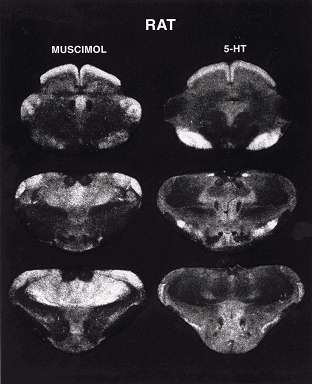
Prints of xray images showing the distribution of the inhibitory neurotransmitter GABA as shown by muscimol; left) and an often excitatory neurotransmitter (serotonin, right) at three levels through the brainstem of the rat. Note that dorsal thalamus is heavily labeled by the inhibitory neurotransmitter but not by the excitatory neurotransmitter. |
Our results support the view that the presence of GABA is common to all of these vertebrate species and thus would be present in humans as well. Further, the amount of GABA was greater in the more `advanced species. And thus, presumably would be greatest in the human brain. Therefore, inhibition by the thalamus is probably a fundamental characteristic of vertebrate species.
The notion that inhibitory gating in the thalamus has been altered during the evolution of the Primates (and therefore human) brain is supported by the results of our study.
One is inclined to think of selection tending
to alter a species, rather than to keep it constant. In point of
fact, selection is probably usually maintaining the constancy of a character
rather than altering it. This seems to be true even at the level of the
biochemistry of the synapses.
Yet, while we know a lot about brain structure and about which brain components are active during the performance of particular functions, we still have no idea about how the brain converts a mass of electrical and chemical signals into what we are familiar with as consciousness and thought patterns. And it is this which is crucial to understand what it is that enables us to be human.
Bruinvels, A. T., Lery, H., Nozulak, J., Palacios, J. M., and Hoyer, D. 5-HT1D binding sites in various species: Similar pharmacological profile in dog, monkey, calf, guinea-pig and human brain membranes. Arch. Pharmacol. 346: 243-248, (1992).
Glendenning, K. K. , Masterton, R. B. Comparative morphometry of mammalian central auditory systems: Variation in nuclei and form of the ascending system. Brain, Behavior & Evolution Feb 1998, Vol 51(2), p 59-89.
Glendenning, K.K.,
Thalamic inhibition in the evolution of human intelligence: Evolutionary
pressure for cortical inhibition . The Mankind Quarterly,
1998, 38:1-18.
Jerison, Harry J. The evolution of intelligence. Sternberg, Robert J., Ed Handbook of intelligence. Cambridge University Press, New York, NY, US; Cambridge University Press, New York, NY, US 2000, p 216-244
Kuhar, M. J. Receptor localization with the microscope. In: Neurotransmitter Receptor Binding, 2nd ed., H. I. Yamamura, S. J. Enna, and M. J. Kuhar, eds., pp. 153-176, Raven Press, New York, (1985).
McKenna, M. C. Toward a phylogenetic classification of Mammalia. In: Phylogeny of the Primates, W. P. Luckett and F. S. Szalay, eds., pp. 21-43, Plenum Press, New York, (1975).
Northcutt, R. G. Evolution of the vertebrate central nervous system: Patterns and processes. Amer. Zool. 24: 701-716, (1984).
Novacek, M. J. Mammalian phylogeny: Shaking the tree. Nature 356: 121-125, (1992).
Rose, J. E., and Woolsey, C. N. Organization of the mammalian thalamus and its relationships to the cerebral cortex. Electroencephalogr. Clin. Neurophysiol. 1: 391-403, (1949).
Smith, G. E.
Some problems relating to the evolution of the brain. Lancet 1: 1-6,
147-153, 221-227, (1910).

Adam Kircher is a National Merit Scholar and attends Maclay School in Tallahassee, Florida. |

Karen Glendenning is a professor of Neuroscience at Florida State University. She has published 49 articles and 3 book chapters. She has given 43 talks at scientific meetings. She was funded by grants from National Institutes of Mental Health for over 25 years to conduct this and other research studies. (Grants NS07726 and DC00197). |